Ide tartoznak az emberi simaizmok is. Sima izmok. Felépítés, funkciók, redukciós mechanizmus. Az izmok osztályozása különféle kritériumok szerint
Az emberi test ingerlékeny szövetei közé tartozik az ideg-, szekréciós és izomszövet. Ez utóbbi azonban más egyedi ingatlan kontraktilitás a speciális fehérjék - miozin, aktin, tropomiozin - mikrofilamentumainak sejtszerkezetében való jelenléte miatt.
Ennek köszönhetően biztosított az ember testtartásának megőrzése, térben való mozgása, a tápbolus elősegítése a gyomor-bél traktus mentén, a vérkeringés és még sok minden más. A szövettani jellemzők, az elvégzett funkciók és az eredet függvényében a simaizom és a harántcsíkolt izomszövetek osztályozása történik, és egyes szerzők a kardiálist a harmadik alfajba is megkülönböztetik. Meg kell azonban érteni, hogy a kontraktilis elemek csak ezeknek a szöveteknek az alapját képezik, és nem működhetnének teljes mértékben a nagy mennyiségű oxigén szállítását biztosító, sűrű véredényhálózat, védő és energetikailag jelentős héj nélkül. sarcolemma, valamint egy támasztó laza formálatlan
barázdált
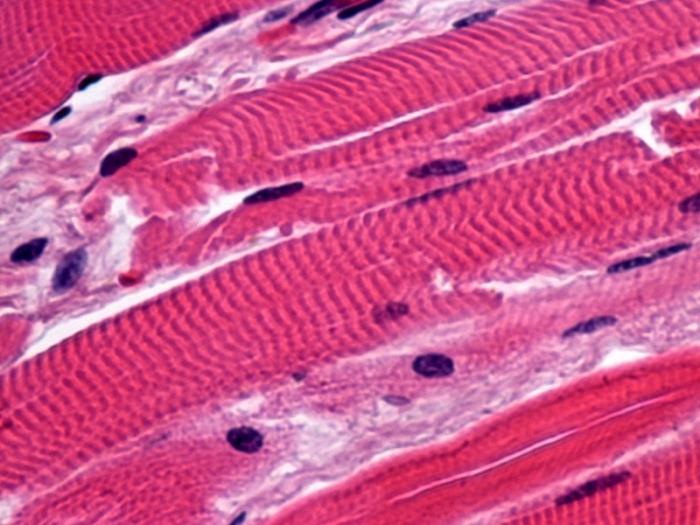
A harántcsíkolt izomszövet alapvetően az összes vázizmot alkotja, és biztosítja az ízületek mozgását és testtartását. Szerkezeti és funkcionális egységük a szarkomer, ezek pedig myosymplasztokból állnak - olyan rostokból, amelyek több egyedi sejt fúziójával jönnek létre a differenciálódási folyamat során. Szövettani preparátumon a harántcsíkolt izomszövetek könnyen megkülönböztethetők többmagvú és harántcsíkolt szerkezetük alapján, ezért kapták nevüket. Másik fontos funkciójuk a hőtermelés, amellyel kapcsolatban a hőmérséklet csökkenésekor hidegrázást tapasztal az ember. Szintén csíkos alkotja a szívizom szerkezetét, csak a szívizomsejteket különböztetik meg a szimplasztok hiánya. Mikroszkóposan egymagvú, orsó alakú sejtek. Feladatuk szerint dolgozókra (többnyire szám szerint), vezetőre és szekrécióra oszthatók. A másodiknak köszönhető, hogy a szív harántcsíkolt izomszövetei az automatizmus tulajdonságával rendelkeznek, azaz képesek önmagukban összehúzódni, ami biztosítja a szív folyamatos működését. A harmadik típusú sejtek a hormonszerű anyagok szintézisének helye, különösen a pitvari nátriuretikus faktor, amely fokozza a diurézist.
Sima
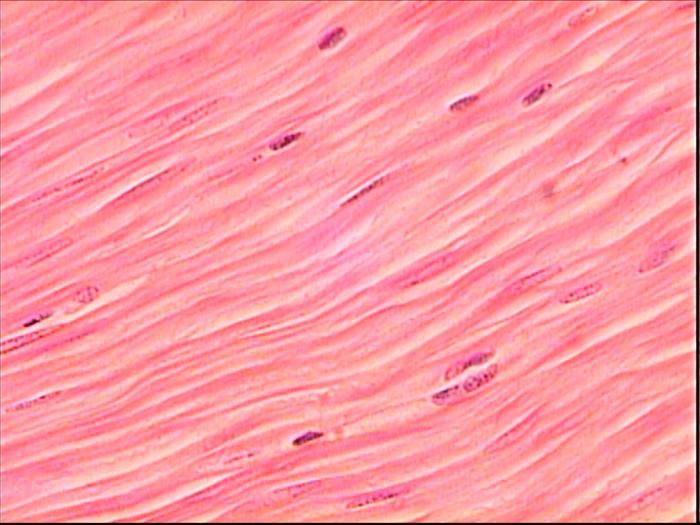
Ha a mozgást biztosító emberi izomszövet harántcsíkolt, akkor a gyomor-bél- és húgyúti traktusok perisztaltikáját, az ér- és hörgők falának összehúzódását a simaizmok biztosítják. Ritmusa, viszonylagos lassúsága, nagyfokú nyújthatósága és regenerációs képessége, valamint autonóm beidegzése jellemzi. Ezek megnyúlt mononukleáris sejtek, csíkozás nélkül, szerkezetükben nagy mennyiségű aktin és kollagén található. Mindegyik ilyen myocitát vékony alapmembrán borítja, és a csoportokat a laza, meg nem formált kötőszövetből származó endomysium borítja.
A simaizmok az üreges szervekben, az erekben és a bőrben találhatók. A simaizomrostok nem rendelkeznek keresztirányú csíkozással. A sejtek megrövidülnek a filamentumok viszonylagos csúszása következtében. Az adenozin-trifoszfát csúszási sebessége és hasadási sebessége 100-1000-szer kisebb, mint az in. Ennek köszönhetően a simaizmok jól alkalmazkodnak a hosszú távú stabil összehúzódáshoz, fáradtság nélkül, kevesebb energiafelhasználással.
Sima izmok vannak szerves része falai egy sor üreges belső szervekés részt vesznek e szervek által ellátott funkciók biztosításában. Különösen szabályozzák a véráramlást a különböző szervekben és szövetekben, a hörgő levegő átjárhatóságát, a folyadékok és a húgyhólyag mozgását (a gyomorban, a belekben, az ureterekben, a vizeletben és az epehólyagban), a méh összehúzódását a szülés során, a pupilla méretét, a bőr könnyítését.
A simaizomsejtek orsó alakúak, 50-400 mikron hosszúak, 2-10 mikron vastagok (5.6. ábra).
A simaizmok akaratlan izmok, azaz. redukciójuk nem függ a makroorganizmus akaratától. A gyomor, a belek, az erek és a bőr motoros aktivitásának jellemzői bizonyos mértékig meghatározzák e szervek simaizmainak élettani jellemzőit.
A simaizom jellemzői
- Automatizmusa van (az intramurális idegrendszer hatása korrekciós)
- Plaszticitás - az a képesség, hogy hosszú ideig megőrizze a hosszt a hang megváltoztatása nélkül
- Funkcionális syncytium - az egyes szálak elkülönülnek, de vannak speciális érintkezési területek - nexusok
- A nyugalmi potenciál értéke 30-50 mV, az akciós potenciál amplitúdója kisebb, mint a vázizomsejteké
- Minimális "kritikus zóna" (gerjesztés akkor következik be, ha bizonyos minimális számú izomelemet gerjesztenek)
- Az aktin és a miozin kölcsönhatásához a Ca 2+ -ion szükséges, amely kívülről érkezik
- Egyetlen összehúzódás időtartama hosszú
a simaizom jellemzője- lassú ritmikus és hosszú tónusos összehúzódások kimutatására való képességük. A gyomor, a belek, az ureterek és más üreges szervek simaizmainak lassú ritmikus összehúzódásai hozzájárulnak tartalmuk mozgásához. Az üreges szervek sphinctereinek simaizomzatának hosszan tartó tónusos összehúzódása megakadályozza tartalmuk önkényes kibocsátását. Az erek falának simaizomzata is állandó tónusos összehúzódás állapotában van, és befolyásolja a vérnyomás szintjét és a szervezet vérellátását.
A simaizomzat egyik fontos tulajdonsága az miszticizmus, azok. a nyújtás vagy deformáció okozta forma megtartásának képessége. A simaizomzat magas plaszticitása nagy jelentőséggel bír a szervek normális működésében. Például a húgyhólyag plaszticitása lehetővé teszi, hogy vizelettel telve megakadályozzuk a nyomásnövekedést a vizeletürítési folyamat megzavarása nélkül.
A simaizmok túlzott nyújtása összehúzódást okoz. Ez a sejtmembránok nyúlásuk okozta depolarizációja következtében következik be, azaz. simaizmok rendelkeznek automatizmus.
A nyújtás okozta összehúzódás fontos szerepet játszik a vérerek tónusának autoregulációjában, a gyomor-bél traktus tartalmának mozgásában és egyéb folyamatokban.
Rizs. 1. A. Vázizomrost, szívizomsejt, simaizomsejt. B. Vázizom szarkomer. B. A simaizom szerkezete. D. A vázizom és a szívizom mechanogramja.
A simaizom automatizmusa a speciális pacemaker (ritmusbeállító) sejtek jelenlétének köszönhető. Szerkezetükben azonosak más simaizomsejtekkel, de különleges elektrofiziológiai tulajdonságokkal rendelkeznek. Ezekben a sejtekben pacemaker-potenciálok keletkeznek, amelyek kritikus szintre depolarizálják a membránt.
A simaizomsejtek gerjesztése fokozza a kalciumionok sejtbe jutását és ezen ionok felszabadulását a szarkoplazmatikus retikulumból. A szarkoplazmában a kalciumionok koncentrációjának növekedése következtében a kontraktilis struktúrák aktiválódnak, de a sima rostokban aktiválódásuk mechanizmusa eltér a harántcsíkolt izmok aktiválódási mechanizmusától. Egy sima sejtben a kalcium kölcsönhatásba lép a kalmodulin fehérjével, amely aktiválja a miozin könnyű láncait. A protofibrillumokban az aktin aktív központjaihoz kapcsolódnak, és "ütést" okoznak. A simaizom passzívan ellazul.
A simaizmok önkéntelenek, és nem függenek az állat akaratától.
A simaizmok élettani tulajdonságai és jellemzői
A simaizmoknak, akárcsak a vázizmoknak, van ingerlékenységük, vezetőképességük és összehúzódásuk. A vázizmokkal ellentétben, amelyek rugalmasak, a simaizmoknak plaszticitásuk van - az a képesség, hogy hosszú ideig fenntartsák a nyújtás során nekik adott hosszt anélkül, hogy növelnék a feszültséget. Ez a tulajdonság fontos az élelmiszerek gyomorban való lerakódásához vagy a folyadékok lerakódásához az epehólyagban és a hólyagban.
A simaizomsejtek ingerlékenységének jellemzői bizonyos mértékig a nyugalmi membránon átívelő alacsony potenciálkülönbséggel járnak együtt (E 0 = (-30) - (-70) mV). A sima myociták lehetnek automatikusak, és spontán akciós potenciált generálnak. Az ilyen sejtek - a simaizom-összehúzódás pacemakerei a bél, a vénás és a nyirokerek falában találhatók.

Rizs. 2. Építsen simán izomsejt(A. Guyton, J. Hall, 2006)
A sima myocytákban az AP időtartama elérheti a tízezredmásodperceket is, mivel bennük az AP elsősorban az intercelluláris folyadékból lassú kalciumcsatornákon keresztül a szarkoplazmába jutó Ca 2+ -ionok miatt alakul ki.
Az AP vezetés sebessége a sima myocyták membránja mentén alacsony - 2-10 cm/s. A vázizmokkal ellentétben a gerjesztés átvihető egyik sima myocytáról a közelben lévő többire. Ez az átvitel az alacsony ellenállású simaizomsejtek közötti kapcsolatok jelenléte miatt következik be. elektromos áram valamint a Ca 2+ -ionok és más molekulák közötti csere biztosítása a sejtek között. Ennek eredményeként a simaizom a funkcionális syncytium tulajdonságait mutatja.
A simaizomsejtek összehúzódását hosszú látens periódus (0,25-1,00 s) és hosszú időtartam (legfeljebb 1 perc) jellemzi egyetlen összehúzódás. A simaizmok kis összehúzódási erőt fejtenek ki, de képesek sokáig tónusos összehúzódásban maradni anélkül, hogy kimerültség alakulna ki. Ez annak köszönhető, hogy a simaizomzat 100-500-szor kevesebb energiát fogyaszt, mint a vázizomzat a tónusos összehúzódás fenntartásához. Ezért a simaizom által elfogyasztott ATP-tartalékoknak még az összehúzódás során is van idejük helyreállni, és egyes testszerkezetek simaizomzata szinte folyamatosan tónusos összehúzódás állapotában van. A simaizom abszolút ereje körülbelül 1 kg/cm 2 .
A simaizom összehúzódásának mechanizmusa
A simaizomsejtek legfontosabb jellemzője, hogy számos inger hatására izgalomba kerülnek. természetes körülmények között csak a felé érkező idegimpulzus indítja be. A simaizom összehúzódását okozhatja mind az idegimpulzusok hatása, mind a hormonok, neurotranszmitterek, prosztaglandinok, egyes metabolitok hatása, valamint fizikai tényezők, például nyújtás hatása. Ezenkívül a sima myocyták gerjesztése és összehúzódása spontán módon is bekövetkezhet - az automatizálás miatt.
A simaizmok azon képessége, hogy összehúzódással reagáljanak különböző tényezők hatására, jelentős nehézségeket okoz ezen izmok tónusának megsértésének korrigálásában. orvosi gyakorlat. Ez látható a bronchiális asztma, az artériás magas vérnyomás, a görcsös vastagbélgyulladás és más olyan betegségek, amelyek a simaizmok összehúzódási aktivitásának korrekcióját igénylő kezelési nehézségekre utalnak.
A simaizom-összehúzódás molekuláris mechanizmusa is számos eltérést mutat a vázizom-összehúzódás mechanizmusától. Az aktin és a miozin filamentumok a simaizomsejtekben kevésbé rendezettek, mint a vázsejtekben, ezért a simaizomzatban nincsenek keresztirányú csíkok. A simaizom aktinszálaiban nincs troponin fehérje, és az aktin központok mindig nyitottak a miozinfejekkel való kölcsönhatásra. Ugyanakkor a miozinfejek nyugalmi állapotban nincsenek energiával ellátva. Az aktin és a miozin kölcsönhatása érdekében a miozinfejeket foszforilálni kell, és több energiát kell adni nekik. Az aktin és a miozin kölcsönhatása a miozinfejek forgásával jár együtt, melynek során az aktin filamentumok a miozin filamentumok közé húzódnak és a sima myocyta összehúzódása következik be.
A miozinfejek foszforilációja a miozin könnyűlánc kináz enzim részvételével, a defoszforiláció pedig foszfatáz segítségével történik. Ha a miozin-foszfatáz aktivitása túlsúlyban van a kináz aktivitásával szemben, akkor a miozinfejek defoszforilálódnak, a miozin és az aktin közötti kapcsolat megszakad, az izom ellazul.
Ezért a zökkenőmentes miocita-összehúzódás érdekében növelni kell a miozin könnyű lánc kináz aktivitását. Tevékenységét a szarkoplazmában lévő Ca 2+ -ionok szintje szabályozza. A neurotranszmitterek (acetilkolin, noradrsnalin) vagy hormonok (vazopresszin, oxitocin, adrenalin) stimulálják specifikus receptorukat, ami a G-fehérje disszociációját okozza, melynek a-alegysége tovább aktiválja a foszfolipáz C enzimet.sejtmembránok. Az IPG az endoplazmatikus retikulumba diffundál, és receptoraival kölcsönhatásba lépve kalciumcsatornák megnyílását és Ca 2+ -ionok felszabadulását idézi elő a depóból a citoplazmába. A citoplazma Ca 2+ -ion-tartalmának növekedése kulcsfontosságú esemény a sima myocyta-összehúzódás megindításában. A szarkoplazmában a Ca 2+ -ionok tartalmának növekedése is az extracelluláris közegből a myocytákba való bejutásának köszönhető (3. ábra).
A Ca 2+ ionok komplexet képeznek a kalmodulin fehérjével, a Ca 2+ -kalmodulin komplex pedig növeli a miozin könnyű láncok kináz aktivitását.
A simaizom-összehúzódás kialakulásához vezető folyamatok sorrendje a következőképpen írható le: Ca 2+ ionok bejutása a szarkoplazmába - kalmodulin aktiválása (a 4Ca 2 -kalmodulin komplex képződésével) - miozin könnyű lánc kináz aktiválása - miozinfejek foszforilációja - a miozinfejek aktinhoz való kötődése és a fejek forgatása, melynek során az aktinszálak behúzódnak a miozin filamentumok közé - összehúzódás.

Rizs. 3. ábra Ca 2+ ionok simaizomsejt szarkoplazmájába való bejutásának útjai (a) és eltávolításuk a szarkoplazmából (b)
A simaizom relaxációhoz szükséges feltételek:
- a szarkoplazma Ca 2+ iontartalmának csökkenése (legfeljebb 10-7 M/l vagy kevesebb);
- a 4Ca 2+ -kalmodulin komplex szétesése, ami a miozin könnyű lánc kináz aktivitásának csökkenéséhez vezet - a miozinfejek defoszforilációja foszfatáz hatására, ami az aktin és a miozin filamentumok kötéseinek megszakadásához vezet.
Ilyen körülmények között a rugalmas erők a simaizomrost eredeti hosszának viszonylag lassú helyreállítását és ellazulását okozzák.
Sima izmok
Főbb pontok A simaizmok egyetlen orsósejtekből állnak.
Általában a simaizmok akaratlan összehúzódásokat hajtanak végre.
A simaizmok az izmos üreges szervek falának fontos részét képezik.
A harántcsíkolt izmokkal ellentétben a simaizom-miofilamentumok nem jól szervezettek, és nincsenek szarkomereik.
A simaizmokkal lehet edzeni speciális gyakorlatok hatékonyságuk és eredményességük javítása érdekében.
Az agy simaizomzatának elsődleges szabályozó elemei az autonóm idegrendszer idegrostjai.
A simaizmok a vér- és nyirokerek falának fő alkotóelemei.
Az erek tónusa határozza meg a véráramlás sebességét és nagyságát.
 A simaizom alakja és elhelyezkedése[szerkesztés]
A simaizom alakja és elhelyezkedése[szerkesztés]
A simaizomsejteket többek között az akaratlan összehúzódások jellemzik. A simaizmok önkéntelenül is képesek összehúzódni, így nincs szükségük akaratlagos kontrollra. Ennek megfelelően elsősorban az emberi test következő szerveiben helyezkednek el.
A simaizmok az izmos üreges szervek falának fontos részét képezik. Ezek közé tartozik az emésztőrendszer és a húgyúti rendszer legtöbb szerve, valamint Légutak.
Az emésztőrendszer a fejből (szájüreg és garat) és az emésztőcső törzsrészeiből (nyelőcső, gyomor és belek), valamint a kapcsolódó külső elválasztású mirigyekből (nyálmirigyek, hasnyálmirigy és máj) áll.
Az emésztőrendszer törzsrészében (csatorna) három simaizomréteg található, amelyek a bélnyálkahártya perisztaltikájáért és mozgásáért felelősek. Egyes területeken különösen hangsúlyos a gyűrűs izomzat, amely a záróizmokat vagy a pylorust és a belső anális záróizmot alkotja.
A törzstől eltérően az emésztőrendszer feje sima izmok helyett harántcsíkolt izmokat tartalmaz.
Az emésztőrendszer falaihoz hasonlóan a húgyúti falakban is három simaizomréteg található, amelyek közelebbről megvizsgálva egyetlen spirális réteget alkotnak.
Mind a férfi, mind a női nemi szervek nagyszámú sima izmok.
A simaizmok nagy mennyiségben találhatók a légcsőben és a hörgőkben - az alsó légutak fontos részein. A simaizomsejtek a légutak átmérőjének változtatásával külső tényezők függvényében szabályozzák a beáramló levegő mennyiségét.
A simaizmok a vér- és nyirokerek részét képezik. Mennyiség simán izomszövet az ér átmérőjével arányos és azonos méretű artériák több simaizmot tartalmaznak, mint a vénák és a nyirokerek. Az erek elágazásával a simaizom abszolút száma is csökken, míg az arteriolákban az átmérőjükhöz viszonyítva a simaizomszövet tartalom maximális. A kapillárisokban nincs simaizomszövet.
A simaizomszövet egyes nem izmos szervekben is megtalálható.
A szemgolyóban a simaizomzat felelős a pupilla tágulásáért és összehúzódásáért, valamint a lencsekapszula feszültségéért. Ez lehetővé teszi, hogy módosítsa a fénytörő erejét és a retinára eső fény mennyiségét. Ez a mechanizmus fontos a közeli és távoli látás és a mélységérzékelés szempontjából.
A hidegre vagy félelemre adott reakcióként a haj felemelése a hajat felemelő izmok (shsh. arrectores pilorum) által történik. Amikor összehúzódnak, a bőr "libabőr" ("libabőr") megjelenését ölti. A szervezetben is (például a külső elválasztású mirigyekben vagy a petefészekben az ovuláció során) vannak olyan sejtek, amelyek kereszteződnek izom- és kötőszöveti sejtek (az úgynevezett myofibroblasztok) vagy hámsejtek (mioepitheliociták) között (Drenckhahn, 2003).
"Figyelem" Ne feledje: Meg kell említeni, hogy egyes izmok, amelyek többé-kevésbé akaratlan összehúzódásokat hajtanak végre, csíkosak. Ezek közé tartoznak a következő izmok.
A rekeszizom a fő légzőizom.
Szívizom.
A szájfenék, a szájpadlás és a garat izmai.
A gége izmai.
Mimikai izmok.
A medencefenék izmai és a külső záróizom (anális és húgycső záróizom).
A simaizom szerkezete
A harántcsíkolt izmokhoz hasonlóan a simaizom összehúzódása a miofilamentumok - vékony aktin és vastag miozin filamentumok (II. típusú miozin) - kölcsönös csúszása miatt következik be. A simaizomsejtek háromszor kevesebb miozint tartalmaznak, mint a harántcsíkolt sejtek. A vázizmokkal ellentétben a simaizomsejtekben a miozin filamentumok véletlenszerűen vannak elrendezve, és nem képeznek szarkomereket és myofibrillumot, ami a „simaizom” kifejezés megjelenésének oka volt. A vékony aktin filamentumok, mint már említettük, a citoplazmában lévő sűrű testekhez vagy a sejtmembránban lévő horgony plakkokhoz kapcsolódnak. Ezek a formációk a Z-vonal analógjai a harántcsíkolt izmokban. A myofilamentumok kaotikus elrendeződése mellett a simaizom kontraktilis apparátusa ultrastrukturálisan és biokémiailag is különbözik a harántcsíkolt izmoktól. Az egyik fontos szerkezeti különbség a Ca2+ csatornák aktivitása és a sebességet befolyásoló ATPáz miozin enzim izomösszehúzódások. A gyengén fejlett szarkoplazmatikus retikulum csak kis mennyiségű Ca2+ ion lerakódását teszi lehetővé, az izomösszehúzódáshoz szükséges ionok többsége pedig a sejtközi térből érkezik a sejt gerjesztésekor. Emiatt a simaizomszövetben egyrészt a Ca2+-ionok lassú árama, másrészt a miozin ATPáz aktivitása alacsonyabb (10-100-szor alacsonyabb, mint a harántcsíkolt izmokban). Így a simaizom-összehúzódásokat alacsony sebesség, de hosszabb időtartam jellemzi (Widmaier et al., 2008).
Összehúzódás
A simaizom izomösszehúzódásainak jellege, folyamata és szabályozása jelentősen eltér a harántcsíkolt izomzatokétól. Ez utóbbiak gyors összehúzódásra képesek és gyorsan elfáradnak, míg a simaizomzatot viszonylag lassú összehúzódás jellemzi, de nagyobb az állóképességük. Ennek oka a simaizomsejtek speciális ultrastruktúrája, valamint a myofilamentumok molekuláris szerkezete. Ezenkívül a simaizomsejtek sejtmembránjának depolarizációját (és ezáltal a Ca2+ ionok bejutását és az ebből eredő összehúzódást) számos tényező okozza, míg a vázizom membránjának depolarizációját az acetilkolin (ACC) neurotranszmitter szabályozza. ) a motoros neuronok választják ki.
Ez utóbbi tulajdonságot tartják a simaizmok akaratlan összehúzódásának okának. Néha azzal érvelnek, hogy a simaizmok egyáltalán nem képesek akaratlagos összehúzódásokra, de ez nem mindig van így. Helyesebb azt állítani, hogy a simaizmoknak nincs szükségük önkéntes kontrollra, hiszen az agytörzsben a vérkeringésért, az emésztőrendszer működéséért stb. felelős központok tudatos kontroll nélkül működnek. Ez igaz a szív és a légzés működését szabályozó idegközpontokra is, bár a szívizom és a légzőizmok (főleg a rekeszizom) harántcsíkolt.
Meg kell jegyezni, hogy a vázizom szabályozása csak részben önkéntes. Az egyetlen izom, amely egyenként (és néhányan csak edzés közben) valódi akaratlagos mozdulatokat hajt végre, a kéz izmai. Az összes többi mozgás sok izom (izom-szinergisták) összetett tudattalan interakciói eredményeként jön létre, mivel a test statikus erői ilyenkor mindig megváltoznak. A kar egyszerű hajlítása a könyöknél megköveteli a kéz, a csukló és a könyök hajlítóinak megfeszítését. Ugyanakkor az antagonista izmoknak is egyszerre kell ellazulniuk (pl. a megfelelő ízületek összes feszítője triceps váll). Ezenkívül a vállöv különböző izmai aktiválódnak, amelyek stabilizálják a lapockot a testhez képest, valamint különféle izmok, amelyek stabilizálják a testet, csípőizületés az alsó végtagok a test statikus (tartás) fenntartása érdekében. Ez a példa azt mutatja, hogy egy izom akaratlagos feszültsége, nem beszélve az egyes izomrostokról, lehetetlen, csakúgy, mint az egyes simaizmok akaratlagos feszültsége. Csak az izomrendszer aktiválása lehetséges, nevezetesen a szinergikusok kombinációja egy mozgás végrehajtásához (például a könyökízület hajlítása).
A fentiek ismeretében az olvasó nem csodálkozhat azon, hogy a simaizomrendszer önkéntes aktiválása továbbra is lehetséges. Például a biofeedback módszerek vagy az edzés eredményeként végzett relaxáció segítségével viszonylag könnyen uralható az izomtevékenység. a szív-érrendszer. Ezeket a módszereket gyakran alkalmazzák a migrén és a magas vérnyomás kezelésére.
Emellett kimutatták, hogy a különböző edzettségi szintek nemcsak a vázizmok intra- és intermuszkuláris koordinációját befolyásolják, hanem a szív- és érrendszer simaizmainak (vérnyomás) vagy az exokrin mirigyek (verejték) aktivitását is, ami jelentősen megváltoztathatja. a sport hatása alatt.edzések.
Végezetül meg kell jegyezni, hogy a simaizom egy bizonyos határig továbbra is önkéntes ellenőrzés alatt áll. Ebből következik egy kézenfekvő, a sportban régóta ismert következtetés: a simaizom speciális technikákkal edzhető, és növelhető teljesítményük és hatékonyságuk. Az egyszerű állóképességi edzés néhány hét után jelentősen növelheti a munka hatékonyságát, például a szív- és érrendszert. Hasonló eredmények ismertek a verejtékmirigyeknél is: az állóképességi gyakorlatok után az edzett emberek hamarabb kezdenek izzadni, mint a nem edzettek.
A simaizom-összehúzódás élettana
A simaizomzat élettani funkcióinak különbségei meghatározzák azok biokémiai és anatómiai jellemzőit. A legfontosabbak a következő jellemzők (Widmaier et al., 2008).
Összehúzódási sebesség- a simaizomsejtek összehúzódása a többi izomszövethez hasonlóan a citoplazmában a Ca2+-ionok koncentrációjának növekedésén alapul. A harántcsíkolt izmokkal ellentétben bennük a szarkoplazmatikus retikulum rosszul fejlett, ezért függővé válnak az ionok sejtbe jutásától. Az ionok bejövő árama sokkal lassabb, mint a szarkoplazmatikus retikulumból a Ca2+ felszabadulási folyamata, ráadásul a miozin enzimek aktivitása a simaizomsejtekben 10-100-szor alacsonyabb, mint a harántcsíkolt sejtekben. Ez a két tényező határozza meg a simaizom-összehúzódások viszonylag alacsony arányát.
Kitartás- a simaizom ellazulása az aktin és a miozin filamentumok közötti keresztkötések felszakadása után következik be. Ehhez a miozinfej defoszforilációja szükséges, mivel csak foszforilált állapotban képes az aktinhoz kötődni. Amikor egy simaizomsejtet gerjesztenek, a miozinfejek foszforilációs és defoszforilációs folyamatai folyamatosan zajlanak, és a foszforiláció sebessége magasabb, mint a defoszforiláció sebessége. A Ca2+ koncentrációjának hosszan tartó növekedésével az aktinhoz már kapcsolódó miozinfejek defoszforilációs folyamatai aktiválódnak. Így a simaizomsejtek több órán keresztül képesek statikus összehúzódást fenntartani kifejezett fáradtság és nagy mennyiségű energia fogyasztása nélkül. Ez a vázizom-rigor mortishoz hasonlítható, azonban in vivo fordul elő, és jellemző a simaizom-záróizomra (pl. hólyagkivezető záróizom).
Fiziológiai hiányosság Ultrastruktúrájuknak köszönhetően a simaizmok erősebb izometrikus és koncentrikus összehúzódásokat hajtanak végre nagyobb hosszúságtartományban, mint a vázizmok. Példa erre a hólyag vagy a méh erős megnyúlása a terhesség végén (ezekben az esetekben az izmok 8-szor nyúlhatnak).
Összehúzódás ellenőrzése
A vázizmokkal ellentétben a simaizom és a szívizmok képesek spontán depolarizációra és összehúzódásra, függetlenül az idegingerektől. A szívben a spontán depolarizáció folyamatai a szerv egyik funkciója, külső vezérlőrendszerrel rendelkeznek, és egészséges szívben csak specifikus izomsejtek csoportja (a sinus és az atrioventrikuláris csomópontok sejtjei) hajtja végre. A simaizomzatban számos tényező okozhatja és befolyásolhatja ezeket a folyamatokat.
vegetativ idegrendszer
A simaizom elsődleges irányító szerve az autonóm idegrendszer (ANS, autonóm idegrendszer) központjai. Filogenetikailag az idegrendszer egy nagyon régi része, túlnyomórészt az agytörzsben található, és a hipotalamusz irányítja. Az ANS részt vesz a homeosztázis fontos paramétereinek fenntartásában és a változó környezeti feltételekhez való alkalmazkodásban, pl. a fizikai aktivitás: fokozott értónus, a légutak tágulása, a bélmotilitás csökkenése stb. az agykéreg közvetlen irányítása nélkül. Háromféle ANS létezik.
Az intraintesztinális vagy enterális, autonóm idegrendszer (ENS - enterális, vagy intramurális, idegrendszer) az idegsejtek felhalmozódása a bélfalban. Egyes simaizom- vagy szívsejtekhez hasonlóan képesek spontán akciós potenciált generálni, és a bélizmok ritmikus hullámzó összehúzódását (perisztaltikát) okozni. Feltehetően az ENS az egyetlen olyan része az idegrendszernek, amely valódi autonómiával rendelkezik, és nem igényli az agy irányítását, amelynek tevékenységét csak az ANS többi központja modulálja. A bélperisztaltika a szervezetből való eltávolítása után is folytatódik, amíg az elektrolit-, oxigén- és tápanyagellátás teljesen ki nem merül.
Szimpatikus idegrendszer - közös funkció Ennek a rendszernek a célja a belső szervek aktivitásának növelése, amely stresszes helyzetekben (harc, menekülés, szexuális aktivitás és sport) szükséges. A szimpatikus idegrendszer fokozza a szívműködést, az érrendszer tónusát és a vérnyomást, kitágítja a felső légutakat, ezáltal fokozza az oxigénellátást, kitágítja a pupillákat, fokozza a verejtékezést stb. Egyúttal csökkenti a bélműködést, a vizelettermelést és a kiválasztó szervek működését. Azoknak az anyagoknak, amelyeken keresztül a szimpatikus idegrendszer hatással van a simaizomzatra, elsősorban az adrenalint és a noradrenalint (körülbelül 80:20 arányban) tekintjük neurotranszmittereknek.
paraszimpatikus idegrendszer- a szimpatikus idegrendszer antagonistájaként tartják számon, mivel ellenkező hatást fejt ki. Egyébként a két típus közötti kölcsönhatás szinergikusnak tekinthető, hiszen a paraszimpatikus idegrendszer feladata a szimpatikus rendszer által igényelt energia felhalmozása. Nyugalomban és az alvás reaktív fázisában a belső szervek paraszimpatikus hatása érvényesül. Mindkét rendszer szinergikus hatása a nemi érintkezés folyamatának példáján is látható, hiszen az erekció a paraszimpatikus rendszer által irányított folyamat, az ejakuláció pedig a szimpatikus rendszer által. A paraszimpatikus rendszer neurotranszmittere az acetilkolin (ACC).
A simaizom kontraktilitását a fenti neurotranszmittereken kívül számos anyag befolyásolja, például a nitrogén-monoxid és a szerotonin vagy mérgek. növényi eredetű muszkarin (légygalóca), atropin (Belladonna/belladonna), nikotin (dohánynövények) és curare (dél-amerikai liliom).
Ca2+ felszabadulása nyújtáskor[Szerk.]
A simaizomzatban akkor is fellépnek akciós potenciálok, amikor megnyújtják őket. Ezzel egyidejűleg a sejtmembránban megnyílnak a nyújtásra érzékeny Ca2+ csatornák, és a Ca2+ ionok berohannak a sejtbe. Ez a mechanizmus nemcsak a homeosztázis fenntartásához rendkívül fontos, hanem a kólikát is okozza. Ebben az esetben az üreges szervek (ureterek, epeutak, belek stb.) simaizmait a fogkő megfeszíti, ami reflexösszehúzódásukhoz vezet. A fogkő jelenléte megzavarja e szervek mozgékonyságát, az izomösszehúzódás nem teszi lehetővé a fogkő előrehaladását, ami újabb összehúzódásokat (időszakos görcsöket) okoz.
A simaizmok élettana
 A simaizom számos orsósejtrétegből áll. A simaizom számos szervben működik (gyomor, belek, epehólyag, hólyag, méh, hörgők, szem stb.), valamint az erekben, ahol fontos szerepük van a vérkeringés szabályozásában. A simaizom egy speciális F-aktin-tropomiozin és miozin II filamentumot tartalmaz, de kevés troponint és miofibrillumot. Ezenkívül a simaizmokban nincs fejlett mikrotubulus- és szarkomérrendszer (nem csíkosak). Innen a név - sima
A simaizom számos orsósejtrétegből áll. A simaizom számos szervben működik (gyomor, belek, epehólyag, hólyag, méh, hörgők, szem stb.), valamint az erekben, ahol fontos szerepük van a vérkeringés szabályozásában. A simaizom egy speciális F-aktin-tropomiozin és miozin II filamentumot tartalmaz, de kevés troponint és miofibrillumot. Ezenkívül a simaizmokban nincs fejlett mikrotubulus- és szarkomérrendszer (nem csíkosak). Innen a név - sima
izomzat.
 A simaizomszálak gyenge kontraktilis apparátust alkotnak, amely hosszanti irányban helyezkedik el a sejtben, és korongos plakkokhoz kapcsolódik (lásd a B modellt), amely a simaizom sejtjei között is mechanikai kommunikációt biztosít. A simaizom sokkal jobban lerövidíthet, mint a harántcsíkolt izom.
A simaizomszálak gyenge kontraktilis apparátust alkotnak, amely hosszanti irányban helyezkedik el a sejtben, és korongos plakkokhoz kapcsolódik (lásd a B modellt), amely a simaizom sejtjei között is mechanikai kommunikációt biztosít. A simaizom sokkal jobban lerövidíthet, mint a harántcsíkolt izom.
Számos szerv (például a belek) simaizomsejtjeinek membránpotenciálja nem állandó, hanem alacsony frekvenciával (3-15 min-1) és amplitúdóval (10-20 mV) ritmikusan változik, így alakul ki. lassú hullámok. Ezek a hullámok akciós potenciál kitöréseket (tüskék) okoznak, ha meghaladnak bizonyos nyugalmi potenciált. Minél tovább marad a lassú hullám a nyugalmi potenciál felett, annál nagyobb számú és gyakoriságú akciós potenciál keletkezik. A csúcs után körülbelül 150 ms-mal viszonylag lassú összehúzódás következik be. A tetanusz meglehetősen alacsony csúcsfrekvenciával fordul elő. Következésképpen a simaizom folyamatosan többé-kevésbé erős összehúzódás (tónus) állapotában van. Egyes szervek simaizomsejtjeinek akciós potenciálja a szív akciós potenciáljához hasonló platóval rendelkezik.
A simaizomnak két típusa van (A). A homogén simaizomszövet sejtjei réskapcsolatokkal kapcsolódnak egymáshoz elektromosan. Az olyan szervekben, mint a gyomor, a belek, az epehólyag, a hólyag, az ureterek, a méh, és bizonyos típusú véredényekben, vagyis ahol ez a fajta simaizomszövet jelen van, az ingerek sejtről sejtre terjednek. Az ingerek autonóm módon jönnek létre a simaizomból (részben a pacemaker sejtek). Más szóval, az inger független a beidegzéstől és sok esetben spontán (miogén tónus). A második típust, a heterogén simaizomszövetet az jellemzi, hogy az intercelluláris interakciók ott főleg az autonóm szerv által továbbított ingerek segítségével valósulnak meg. idegrendszer(neurogén tónus). Ez olyan struktúrákban fordul elő, mint az arteriolák, ondócsövek, írisz, ciliáris test, a hajgyökerek izmai. Mivel ezek között a simaizomsejtek között általában nincsenek hézagok, a stimuláció lokális marad, mint a vázizom motoros egységeiben.
A sima izomtónust a depolarizáció mértéke szabályozza (például nyújtó vagy pacemaker sejteken keresztül), valamint mediátorok (például acetilkolin vagy noradrenalin) és számos hormon (például a méhben - ösztrogén, progeszteron és oxitocin, az erek falában - hisztamin, angiotenzin II, vazopresszin, szerotonin és bradikinin). A tónusnövekedés akkor következik be, ha ezen tényezők bármelyike közvetlenül vagy közvetve 10-6 mol/l fölé emeli a Ca2+ intracelluláris koncentrációját. A Ca2+ beáramlása elsősorban a sejtközi térből történik, de kis mennyiségben az intracelluláris raktárakból is (B1) érkezik. A Ca2+ ionokat a kalmodulin (CM) (B2) köti meg, a Ca2+-CM komplex pedig elősegíti a kontrakciót. Hogyan történik?
Szabályozás a miozin II (B3) szintjén: a Ca2+-CM komplex aktiválja a miozin könnyű lánc kinázt (MLCK), amely adott pozícióban foszforilálja a miozin szabályozó könnyűláncot (RLCK), aminek köszönhetően a miozin fej kölcsönhatásba léphet az aktinnal (B6).
Aktinszintű szabályozás (B4): A Ca2+-CM komplex megköti a caldesmont (CDM) is, amely aztán lehasad az aktin-tropomiozin komplexről, így elérhetővé válik a filamentum siklása számára (B6). Úgy tűnik, hogy a KDM protein-kináz C (PC-C) általi foszforilációja szintén képes a filamentum csúszását indukálni (B5).
A következő tényezők vezetnek a tónus csökkenéséhez: a citoplazmában a Ca2+ koncentrációjának csökkenése 10-6 mol / l alá (B7), a foszfatáz aktivitás (B8), valamint a protein kináz C aktivitása, amikor foszforilálódik. a miozin könnyű lánc másik pozíciója (B9).
A simaizom "hossz-erő" összefüggésének rögzítésekor az izomerő állandó csökkenése látható, miközben az izom hossza állandó marad. Az izomnak ezt a tulajdonságát plaszticitásnak nevezzük.
fontos a simaizom tulajdonságai nagy plaszticitása, vagyis az a képesség, hogy a nyújtás által adott hosszt megtartja anélkül, hogy a feszültség változna. A csekély plaszticitású vázizom és a jól meghatározott plaszticitással rendelkező simaizom közötti különbség könnyen kimutatható, ha először lassan megnyújtjuk, majd a húzóterhelést megszüntetjük. a terhelés eltávolítása után azonnal lerövidül. Ezzel szemben a simaizom a terhelés eltávolítása után addig marad megfeszülve, amíg valamilyen irritáció hatására aktív összehúzódása meg nem történik.
A plaszticitás tulajdonsága nagyon fontos az üreges szervek falának, így a hólyag simaizmainak normál működéséhez: a hólyag falának simaizmainak plaszticitása miatt a benne lévő nyomás viszonylag kis mértékben változik különböző töltési fokozatok.
Izgatottság és izgalom
Sima izmok kevésbé ingerlékenyek, mint a csontvázak: magasabbak az irritációs küszöbük, és hosszabb a kronaxia. A legtöbb simaizomrost akciós potenciálja kis amplitúdójú (körülbelül 60 mV a vázizomrostok 120 mV helyett) és hosszú időtartamú - akár 1-3 másodpercig. A rizs. 151 a méhizom egyetlen rostjának akciós potenciálját mutatja be.
A refrakter periódus az akciós potenciál teljes időtartama alatt, azaz 1-3 másodpercig tart. A gerjesztés vezetési sebessége a különböző szálakban másodpercenként néhány millimétertől néhány centiméterig változik.
Nagyon sok van különféle típusok simaizmok az állatok és az emberek testében. A test üreges szerveinek többsége simaizmokkal van bélelve, amelyek érzékeny szerkezetűek. Az ilyen izmok egyes rostjai nagyon szorosan szomszédosak egymással, és úgy tűnik, hogy morfológiailag egyetlen egészet alkotnak.
Az elektronmikroszkópos vizsgálatok azonban kimutatták, hogy az izom-szincicium egyes rostjai között nincs membrán és protoplazma folytonosság: vékony (200-500 Å) rések választják el őket egymástól. A "szinciciális szerkezet" fogalma jelenleg inkább fiziológiai, mint morfológiai.
syncytium- ez egy funkcionális képződmény, amely biztosítja, hogy az akciós potenciálok és a lassú depolarizációs hullámok szabadon terjedjenek egyik szálról a másikra. Az idegvégződések csak kis számú syncytium roston találhatók. A gerjesztés egyik rostról a másikra való akadálytalan terjedése miatt azonban a teljes izom részvétele a reakcióban bekövetkezhet, ha az idegimpulzus kevés izomrosthoz érkezik.
Sima izomösszehúzódás
Egyetlen irritáció nagy ereje esetén simaizom-összehúzódás léphet fel. Ennek az izomnak az egyszeri összehúzódásának látens periódusa sokkal hosszabb, mint egy vázizomé, például egy nyúl bélizomzatában eléri a 0,25-1 másodpercet. Maga a kontrakció időtartama is nagy ( rizs. 152): egy nyúl gyomrában eléri az 5 másodpercet, a béka gyomrában pedig az 1 percet vagy többet. Összehúzódás után különösen lassú a relaxáció. Az összehúzódási hullám a simaizomzaton is nagyon lassan terjed, másodpercenként mindössze 3 cm-rel halad. De a simaizom összehúzódásának ez a lassúsága nagy erejükkel párosul. Így a madarak gyomrának izmai keresztmetszetük 1 cm2-énként 1 kg-ot képesek megemelni.
Sima izomtónus
Az összehúzódás lassúsága miatt egy simaizom még ritka ritmusos ingerek mellett is (békagyomránál percenként 10-12 inger is elég) könnyen átmegy tartós, tartós összehúzódásos állapotba, ami a vázizomzat tetanuszára emlékeztet. . A simaizom ilyen tartós összehúzódása során azonban az energiafelhasználás nagyon kicsi, ami megkülönbözteti ezt a kontrakciót a harántcsíkolt izom tetanuszától.
Az okok, amelyek miatt a simaizmok sokkal lassabban húzódnak össze és ellazulnak, mint a vázizmok, még nem teljesen tisztázottak. Ismeretes, hogy a simaizom miofibrillumai, akárcsak a vázizmoké, miozinból és aktinból állnak. A simaizmokban azonban nincs csíkozás, nincs Z membrán, és sokkal gazdagabbak a szarkoplazmában. Nyilvánvalóan a simaizomhullámok szerkezetének ezen jellemzői határozzák meg a kontraktilis folyamat lassú ütemét. Ez megfelel a relatív alacsony szint simaizom anyagcserét.
Sima izom automatizálás
A simaizmok jellegzetes tulajdonsága, amely megkülönbözteti őket a vázizmoktól, a spontán automatikus tevékenység képessége. Spontán összehúzódások figyelhetők meg a gyomor, a belek, az epehólyag, az ureterek és számos más simaizomszerv simaizmainak vizsgálatakor.
A simaizom automatizálás miogén eredetű. Magukban az izomrostokban rejlik, és a simaizom szervek falában található idegelemek szabályozzák. Az automatizmus miogén természetét a bélfal izomcsíkjain végzett kísérletek igazolták, amelyeket gondos kimetszéssel szabadítottak meg a szomszédos idegfonatokból. Az ilyen csíkok, amelyeket meleg, oxigénnel telített Ringer-Locke oldatba helyeznek, képesek automatikus összehúzódásra. A későbbi szövettani vizsgálat kimutatta az idegsejtek hiányát ezekben az izomcsíkokban.
A simaizomrostokban a membránpotenciál következő spontán oszcillációi különböztethetők meg: 1) lassú depolarizációs hullámok, amelyek ciklusideje körülbelül néhány perc és amplitúdója körülbelül 20 mV; 2) kis gyors potenciál-ingadozások, amelyek megelőzik az akciós potenciálok megjelenését; 3) akciós potenciálok.
A simaizom minden külső hatásra a spontán ritmus frekvenciájának megváltoztatásával reagál, ami az izom összehúzódását és ellazulását eredményezi. A bél simaizomzatának irritációjának hatása a stimuláció gyakoriságának és a spontán ritmus természetes frekvenciájának arányától függ: alacsony tónus esetén - ritka spontán akciós potenciálok esetén - az alkalmazott irritáció növeli a tónust; magas tónus esetén , az ellazulás az irritáció hatására következik be, mivel az impulzusok túlzott növekedése ahhoz vezet, hogy minden következő impulzus a refrakter fázisba esik az előzőtől.
Sima izmok jelen vannak az emésztőcsatorna falában, a hörgőkben, a vér- és nyirokerekben, a hólyagban, a méhben, valamint az íriszben, a ciliáris izomzatban, a bőrben és a mirigyekben. A harántcsíkolt izmokkal ellentétben ezek nem különálló izmok, hanem csak a szervek egy része. A simaizomsejtek megnyúlt, fusiform vagy szalagszerű alakúak, hegyes végekkel. Hosszúságuk emberben általában körülbelül 20 mikron. A legnagyobb hosszúságot (akár 500 mikronig) az emberi terhes méh falában lévő simaizomsejtek érik el. A sejt középső részében egy pálcika alakú mag található, a citoplazmában pedig az egész sejt mentén párhuzamosan futnak egymással a legvékonyabb, teljesen homogén miofibrillumok. Ezért a sejtnek nincs keresztirányú csíkozása. A vastagabb myofibrillumok a sejt külső rétegeiben helyezkednek el. Ezeket határnak nevezik, és egytengelyű kettős törésük van. Elektronmikroszkópban látható, hogy a miofibrillák protofibrillumok kötegei, és keresztirányú csíkozásuk van, amely fénymikroszkópban nem látható. A simaizomsejtek osztódással (mitózissal) regenerálódhatnak. Különféle aktomiozint – tonoaktomiozint tartalmaznak. A simaizomsejtek között ugyanazok a membránkontaktusok vagy nexusok vannak, mint a szívsejtek között, amelyek mentén a gerjesztés és a gátlás átterjed egyik simaizomsejtről a másikra.
A simaizomzatban a gerjesztés lassan terjed.A simaizom összehúzódását erősebb és hosszabb ingerek okozzák, mint a vázizomzatot. Összehúzódásának látens periódusa néhány másodpercig tart. A simaizmok sokkal lassabban húzódnak össze, mint a vázizmok. Így a béka gyomrában a simaizom összehúzódási periódusa 15-20 s. A simaizom-összehúzódások több percig vagy akár órákig is eltarthatnak. A vázizmokkal ellentétben a simaizom-összehúzódások tónusosak. A simaizmok rendkívül alacsony anyag- és energiafelhasználással képesek hosszú ideig tónusos feszültség állapotában lenni. Például az emésztőcsatorna, a hólyag, az epehólyag, a méh és más szervek sphinctereinek simaizomzata több tíz percig és sok óráig jó formában van. A magasabb gerincesek ereinek falának simaizomzata egész életen át jó formában marad.
Közvetlen kapcsolat van az izomban fellépő impulzusok gyakorisága és az izomfeszülés mértéke között. Minél nagyobb a frekvencia, annál nagyobb a tónus egy bizonyos határig a nem egyidejűleg megfeszülő izomrostok feszültségeinek összegzése miatt.
A simaizmok ízletesek – képesek megőrizni hosszukat nyújtáskor anélkül, hogy a feszültség megváltozna, ellentétben a vázizmokkal, amelyek megfeszülnek nyújtáskor.
A vázizmokkal ellentétben sok simaizom automatikus. A lokális reflexmechanizmusok hatására összehúzódnak, mint például a tápcsatornában lévő Meisner és Auerbach plexusok, ill. vegyi anyagok a vérbe jutó, például acetilkolin, noradrenalin és epinefrin. Az idegrendszerből érkező idegimpulzusok hatására a simaizomzat automatikus összehúzódásai felerősödnek vagy gátolódnak. Ezért a vázizmokkal ellentétben vannak speciális gátló idegek, amelyek megállítják az összehúzódást és a simaizom ellazulását okozzák. Egyes simaizmok, amelyekben nagyszámú idegvégződés van, nem rendelkezik automatizmussal, például a pupilla záróizomja, a macska nyelőhártyája.
A simaizmok jelentősen lerövidíthetők, sokkal jobban, mint a vázizmok. Egyetlen stimuláció 45%-os simaizom-összehúzódást okozhat, gyakori stimulációs ritmus mellett a maximális összehúzódás elérheti a 60-75%-ot.
A mezodermából simaizomszövet is fejlődik (a mesenchymából származik); egyedi, erősen megnyúlt orsó alakú sejtekből áll, amelyek mérete jóval kisebb a harántcsíkolt izmok rostjaihoz képest. Hosszúságuk 20-500 µm, szélességük 4-7 µm. Általában ezeknek a sejteknek egy hosszúkás magja van, amely a sejt közepén fekszik. A sejt protoplazmájában számos és nagyon vékony miofibrillum fut hosszirányban, amelyeken nincsenek keresztirányú csíkok, és speciális kezelés nélkül teljesen láthatatlanok. Minden simaizomsejt a legvékonyabb kötőszöveti hüvelybe van öltözve. Ezek a membránok a szomszédos sejtek kapcsolódnak egymáshoz. A vázizomzat szinte teljes hosszában elhelyezkedő harántcsíkolt rostokkal ellentétben minden simaizom komplexumban jelentős számú sejt található egy vonalban.
A simaizomsejtek a testben vagy külön-külön a kötőszövetben találhatók, vagy különböző méretű izomkomplexumokban társulva.
Utóbbi esetben az egyes izomsejteket minden oldalról egy-egy intercelluláris anyag is körülveszi, amelybe a legfinomabb fibrillumok hatolnak be, ezek száma nagyon eltérő lehet. Az intercelluláris anyagban a rugalmas rostok legfinomabb hálózatai is megtalálhatók.
A szervek simaizomsejtjei izomkötegekké egyesülnek. Sok esetben (húgyúti, méh stb.) ezek a kötegek elágaznak és más kötegekkel egyesülnek, változó sűrűségű felületi hálózatokat alkotva. Ha nagyszámú köteg található szorosan, akkor sűrű izommembrán képződik (például a gyomor-bél traktus). A simaizmok vérellátását az ereken keresztül végzik, amelyek nagy kötőszöveti rétegekben haladnak át a kötegek között; kapillárisok behatolnak az egyes kötegek rostjai közé, és azok mentén elágazva sűrű kapilláris hálózatot alkotnak. A simaizomszövet nyirokereket is tartalmaz. A simaizmokat az autonóm idegrendszer rostjai beidegzik. A simaizomsejtek a harántcsíkolt izomrostokkal ellentétben lassú, tartós összehúzódásokat produkálnak. Hosszú ideig és nagy erővel képesek dolgozni. Például a méh izmos falai a szülés során, amelyek órákig tartanak, olyan erőt fejlesztenek ki, amely a harántcsíkolt izmok számára elérhetetlen. A simaizmok tevékenysége általában nem függ akaratunktól (vegetatív beidegzés, lásd alább) - ezek önkéntelenek.
A simaizmok fejlődésükben (filogenezis) ősibbek, mint a harántcsíkolt izmok, és gyakoribbak az állatvilág alsóbb formáiban.
A simaizom osztályozása
A simaizmokat zsigeri (egységes) és több egységre osztják. A zsigeri simaizom minden belső szervben, az emésztőmirigyek csatornáiban, a vér- és nyirokerekben, valamint a bőrben megtalálható. A több egységből álló izmok közé tartozik a ciliáris izom és az írisz izma. A simaizom zsigeri és több egységre való felosztása motoros beidegzésük eltérő sűrűségén alapul. A zsigeri simaizomzatban a motoros idegvégződések kis számú simaizomsejten találhatók. Ennek ellenére az idegvégződésekből származó gerjesztés a köteg összes simaizomsejtjébe továbbítódik a szomszédos miociták - nexusok közötti szoros érintkezés miatt. A nexusok lehetővé teszik az akciós potenciálok és a lassú depolarizációs hullámok terjedését egyik izomsejtről a másikra, így a zsigeri simaizom az idegimpulzus érkezésével egyidejűleg összehúzódik.
A simaizom funkciói és tulajdonságai
Műanyag. A simaizom másik fontos sajátossága a feszültség változékonysága, anélkül, hogy a hosszával szabályos összefüggés lenne. Így ha egy zsigeri simaizom megfeszül, akkor a feszültsége nő, ha viszont az izom nyújtás okozta megnyúlás állapotában tartjuk, akkor a feszültség fokozatosan csökken, esetenként nem csak a nyújtás előtti szintre, hanem ezen szint alatt is. Ezt a tulajdonságot simaizom plaszticitásnak nevezik. Így a simaizom jobban hasonlít egy viszkózus képlékeny tömeghez, mint egy strukturált szövethez, amelynek alacsony a megfelelősége. A simaizmok plaszticitása hozzájárul a belső üreges szervek normál működéséhez.
A gerjesztés és az összehúzódás kapcsolata. A zsigeri simaizomban nehezebb vizsgálni az elektromos és mechanikai megnyilvánulások kapcsolatát, mint a vázizomban vagy a szívizomban, mivel a zsigeri simaizom folyamatos aktivitásban van. Relatív pihenés körülményei között egyetlen AP regisztrálható. Mind a váz-, mind a simaizom összehúzódása az aktin miozinhoz viszonyított csúszásán alapul, ahol a Ca2+ ion trigger funkciót lát el.
A simaizom-összehúzódás mechanizmusának van egy olyan tulajdonsága, amely megkülönbözteti a vázizom-összehúzódás mechanizmusától. Ez a tulajdonság abban rejlik, hogy mielőtt a simaizom miozin megmutathatná ATP-áz aktivitását, foszforilálni kell. A miozin foszforilációja és defoszforilációja a vázizomzatban is megfigyelhető, de abban a foszforiláció folyamata nem szükséges a miozin ATPáz aktivitásának aktiválásához. A simaizom miozin foszforilációja a következőképpen zajlik: a Ca2+ ion kalmodulinnal kombinálódik (a kalmodulin a Ca2+ ion receptorfehérje). A kapott komplex aktiválja a miozin könnyű lánc kináz enzimet, ami viszont katalizálja a miozin foszforilációs folyamatát. Ezután az aktin elcsúszik a miozinhoz képest, amely az összehúzódás alapját képezi. Megjegyzendő, hogy a simaizom-összehúzódás kiindulópontja a Ca2+ ion kalmodulinhoz való kötődése, míg a váz- és szívizomban a Ca2+ troponinhoz való kötődése.
Kémiai érzékenység. A simaizom rendkívül érzékeny a különféle fiziológiailag aktív anyagokra: adrenalinra, noradrenalinra, ACh-ra, hisztaminra stb. Ennek oka a simaizomsejtek membránján található specifikus receptorok jelenléte. Ha adrenalint vagy noradrenalint adunk a bél simaizom készítményéhez, megnő a membránpotenciál, csökken az AP gyakorisága, és az izom ellazul, azaz ugyanaz a hatás figyelhető meg, mint a szimpatikus idegek gerjesztésekor.
A noradrenalin a simaizomsejtek membránjának α- és β-adrenerg receptoraira hat. A noradrenalin és a β-receptorok kölcsönhatása csökkenti az izomtónust az adenilát-cikláz aktiválódása és a ciklikus AMP képződése és az ezt követő intracelluláris Ca2+-kötés fokozódása következtében. A noradrenalin α-receptorokra gyakorolt hatása gátolja a kontrakciót azáltal, hogy fokozza a Ca2+-ionok felszabadulását az izomsejtekből.
Az ACh a noradrenalin hatásával ellentétes hatással van a membránpotenciálra és a bél simaizmainak összehúzódására. Az ACh hozzáadása a bél simaizom készítményéhez csökkenti a membránpotenciált és növeli a spontán AP-k gyakoriságát. Ennek eredményeként nő a tónus és a ritmikus összehúzódások gyakorisága, azaz ugyanaz a hatás figyelhető meg, mint a paraszimpatikus idegek gerjesztésekor. Az ACh depolarizálja a membránt, növeli Na+ és Ca+ permeabilitását.
Egyes szervek simaizomzata különféle hormonokra reagál. Így az állatok méhének simaizomzata az ovuláció közötti időszakokban és a petefészkek eltávolítása során viszonylag ingerlhetetlen. Ösztrusz alatt vagy petefészkektől megfosztott állatoknál, amelyekbe ösztrogént fecskendeztek be, a simaizmok ingerlékenysége megnő. A progeszteron még az ösztrogénnél is jobban növeli a membránpotenciált, de ebben az esetben a méh izmainak elektromos és összehúzó aktivitása gátolt.
A simaizom a belső szervek része. Az összehúzódásnak köszönhetően biztosítják szerveik (emésztőcsatorna, húgyúti rendszer, erek stb.) motoros (motoros) funkcióját. A vázizmokkal ellentétben a simaizmok akaratlanok.
A sima morfo-funkcionális szerkezete izmok. A simaizom fő szerkezeti egysége az izomsejt, amely orsó alakú, és kívülről plazmamembrán borítja. Elektronmikroszkóp alatt számos mélyedés látható a membránban - caveolák, amelyek jelentősen megnövelik az izomsejt teljes felületét. Az unfazed izomsejt szarkolemmája a plazmamembránt, az azt kívülről lefedő alapmembránnal és a szomszédos kollagénrostokkal tartalmazza. A fő intracelluláris elemek: sejtmag, mitokondriumok, lizoszómák, mikrotubulusok, szarkoplazmatikus retikulum és kontraktilis fehérjék.
Az izomsejtek izomkötegeket és izomrétegeket alkotnak. Az intercelluláris tér (100 nm vagy több) elasztikus és kollagén rostokkal, kapillárisokkal, fibroblasztokkal stb. van kitöltve. Egyes területeken a szomszédos sejtek membránjai nagyon szorosan fekszenek (a sejtek közötti rés 2-3 nm). Feltételezhető, hogy ezek a területek (nexus) az intercelluláris kommunikációt, a gerjesztés továbbítását szolgálják. Bebizonyosodott, hogy egyes simaizomzatok nagy számban tartalmaznak nexust (pupilla záróizmok, vékonybél körkörös izmai stb.), míg másokban kevés, vagy egyáltalán nincs (vas deferens, bél hosszanti izmai). A nemdohányzó izomsejtek között köztes, vagy desmotikus kapcsolat is van (membrán megvastagodásával és sejtfolyamatok segítségével). Nyilvánvalóan ezek a kapcsolatok fontosak a sejtek mechanikai összekapcsolásához és a mechanikai erő sejtek általi átviteléhez.
A miozin és aktin protofibrillumok kaotikus eloszlása miatt a simaizomsejtek nem csíkozottak, mint a váz- és szívsejtek. A vázizmokkal ellentétben a simaizmokban nincs T-rendszer, a szarkoplazmatikus retikulum a myoplazma térfogatának mindössze 2-7%-át teszi ki, és nincs kapcsolata a sejt külső környezetével.
A simaizomzat élettani tulajdonságai .
A simaizomsejtek, mint a harántcsíkolt izmok, összehúzódnak az aktin protofibrillumok miozinok közötti csúszása miatt, de a csúszás és az ATP hidrolízis sebessége, így a kontrakció sebessége 100-1000-szer kisebb, mint a harántcsíkolt izmokban. Ennek köszönhetően a simaizmok jól alkalmazkodnak a hosszú távú csúszáshoz, kevés energiával és fáradtság nélkül.
A sima izmokat, figyelembe véve azt a képességet, hogy AP-t generáljanak válaszként a küszöbértékre vagy a szarv feletti stimulációra, hagyományosan fázisos és tónusos izomzatra oszthatók. A fázisos izmok teljes értékű AP-t generálnak, a tónusos izmok - csak lokálisak, bár van egy mechanizmusuk is a teljes értékű potenciál generálására. A tónusos izmok képtelensége az AP-ra a membrán magas kálium-permeabilitásával magyarázható, ami megakadályozza a regeneratív depolarizáció kialakulását.
Az el nem söpört izmok simaizomsejtjeinek membránpotenciáljának értéke -50 és -60 mV között változik. Mint a többi izomban, beleértve idegsejtek, kialakulásában főleg a +, Na +, Cl- vesznek részt. A tápcsatorna, a méh és egyes erek simaizomsejtjeiben a membránpotenciál instabil, lassú depolarizációs hullámok formájában spontán fluktuációk figyelhetők meg, amelyek tetején AP kisülések jelenhetnek meg. Az AP időtartama a simaizomzatban 20-25 ms-tól 1 s-ig vagy még tovább terjed (például a hólyag izmaiban), pl. hosszabb, mint a vázizmok AP időtartama. A simaizom AP mechanizmusában a Na + mellett nagy szerepet Ca2+-t játszik.
Spontán miogén aktivitás. A vázizmokkal ellentétben a gyomor, a belek, a méh és az ureterek simaizomzata spontán miogén aktivitással rendelkezik, pl. spontán tetanohyodibne összehúzódások alakulnak ki. Ezeket az izmok izolálása és az intrafuzális idegfonatok farmakológiai leállítása mellett tárolják. Tehát a PD magukban a simaizmokban fordul elő, és nem az idegimpulzusok izmokhoz való átvitelének köszönhető.
Ez a spontán aktivitás miogén eredetű, és olyan izomsejtekben fordul elő, amelyek pacemakerként működnek. Ezekben a sejtekben a lokális potenciál elér egy kritikus szintet és átalakul AP-vé. De a membrán repolarizációja után spontán módon új lokális potenciál keletkezik, ami újabb AP-t okoz, stb. A nexuson keresztül a szomszédos izomsejtekhez 0,05-0,1 m/s sebességgel terjedő AP a teljes izmot lefedi, ennek összehúzódását okozva. Például a gyomor perisztaltikus összehúzódásai 1 percenként 3-szor fordulnak elő, a vastagbél szegmentális és ingamozgásai - 20-szor 1 percenként a felső szakaszokban és 5-10 percenként - az alsóban. Így e belső szervek simaizomrostjai automatizmussal rendelkeznek, ami abban nyilvánul meg, hogy külső ingerek hiányában képesek ritmikusan összehúzódni.
Mi az oka a potenciál megjelenésének a pacemaker simaizomsejtjeiben? Nyilvánvalóan a kálium csökkenése és a membrán nátrium- és kalcium-permeabilitásának növekedése miatt következik be. Ami a lassú depolarizációs hullámok rendszeres előfordulását illeti, amelyek a legkifejezettebbek a gyomor-bél traktus izmaiban, nincs megbízható adat az ionos eredetükről. Lehetséges, hogy bizonyos szerepet játszik a káliumáram kezdeti inaktiváló komponensének csökkenése az izomsejtek depolarizációja során a megfelelő kálium-ioncsatornák inaktiválódása miatt.
A simaizmok rugalmassága és nyújthatósága. A vázizmokkal ellentétben plasztikus, rugalmas szerkezetként simák, ha megfeszítik magukat. A plaszticitásnak köszönhetően a simaizom összehúzott és feszített állapotban is teljesen ellazítható. Például a gyomor vagy a hólyag falának simaizmainak plaszticitása, mivel ezek a szervek megtelnek, megakadályozza az intracavitaris nyomás növekedését. A túlzott nyújtás gyakran az összehúzódás stimulálásához vezet, ami a pacemaker sejtek depolarizációja miatt következik be, ami az izom megfeszítésekor következik be, és az AP gyakoriságának növekedésével és ennek következtében az összehúzódás növekedésével jár. Az erek alaptónusának önszabályozásában nagy szerepe van a nyújtási folyamatot aktiváló összehúzódásnak.
simaizom-összehúzódás mechanizmusa. A simaizom, valamint a vázizom összehúzódásának előfeltétele a Ca2 + koncentrációjának növekedése a myoplazmában (10v-5 M-ig). Úgy gondolják, hogy a kontrakciós folyamatot főként az extracelluláris Ca2 + aktiválja, amely feszültségfüggő Ca2 + csatornákon keresztül jut be az izomsejtekbe.
A simaizom neuromuszkuláris transzmissziójának sajátossága, hogy a beidegzést az autonóm idegrendszer végzi, és serkentő és gátló hatása is lehet. Típusuk szerint megkülönböztetünk kolinerg (mediátor acetilkolin) és adrenerg (mediátor noradrenalin) mediátorokat. Előbbiek általában az emésztőrendszer, utóbbiak az erek izmaiban találhatók.
Ugyanaz a mediátor egyes szinapszisokban serkentő, másokban gátló hatású lehet (a citoreceptorok tulajdonságaitól függően). Az adrenoreceptorok a- és B-re oszlanak. Az a-adrenerg receptorokra ható noradrenalin összehúzza az ereket és gátolja az emésztőrendszer mozgékonyságát, a B-adrenerg receptorokra hatva serkenti a szívműködést és kitágítja egyes szervek ereit, ellazítja a hörgők izmait. . Leírt neuromuszkuláris. ny transzfer a simaizmokban a segítségért és egyéb közvetítőkért.
Egy serkentő mediátor hatására a simaizomsejtek depolarizációja következik be, ami serkentő szinaptikus potenciál (SSP) formájában nyilvánul meg. Amikor eléri a kritikus szintet, PD lép fel. Ez akkor történik, amikor több impulzus érkezik egymás után az idegvégződéshez. Az ISGI megjelenése a posztszinaptikus membrán Na +, Ca2 + és SI permeabilitásának növekedésének a következménye.
A gátló neurotranszmitter a posztszinaptikus membrán hiperpolarizációját okozza, ami a gátló szinaptikus potenciálban (GSP) nyilvánul meg. A hiperpolarizáció a membránpermeabilitás növekedésén alapul, főleg a K+ esetében. A gátló mediátor szerepét az acetilkolin által gerjesztett simaizomzatban (például a bélizmokban, hörgőkben) a noradrenalin, az acetilkolin pedig azokban a simaizomzatokban, amelyeknél a noradrenalin serkentő közvetítő (például a hólyagizmok).
Klinikai és fiziológiai szempont. Egyes betegségekben a vázizmok beidegzésének zavara esetén passzív nyújtásuk vagy elmozdulásuk tónusuk reflexszerű növekedésével jár, pl. nyúlással szembeni ellenállás (spaszticitás vagy merevség).
Keringési zavarok, valamint bizonyos anyagcseretermékek (tej- és foszforsav), mérgező anyagok, alkohol, fáradtság, izomhőmérséklet csökkenés (például hosszan tartó hideg vízben úszás) esetén kontraktúra léphet fel. hosszan tartó aktív izomösszehúzódás után. Minél jobban zavart az izomműködés, annál kifejezettebb a kontraktúra utóhatása (például a rágóizmok összehúzódása a maxillofacialis régió patológiájában). Mi a kontraktúra eredete? Úgy gondolják, hogy a kontraktúra az ATP koncentrációjának csökkenése miatt keletkezett az izomban, ami állandó kapcsolat kialakulásához vezetett a keresztirányú hidak és az aktin protofibrillumok között. Ebben az esetben az izom elveszíti rugalmasságát és megkeményedik. A kontraktúra alábbhagy, az izom ellazul, amikor az ATP-koncentráció eléri a normál szintet.
Az olyan betegségekben, mint a myotonia, az izomsejtek membránja olyan könnyen izgat, hogy még enyhe irritáció is (például tűelektróda bevezetése az elektromiográfia során) izomimpulzusok kisülését okozza. A spontán AP-t (fibrillációs potenciál) az izomdenervációt követő első szakaszban is rögzítik (amíg az inaktivitás az izomsorvadáshoz vezet).






